Despite reductions in use and production worldwide, the risk of asbestos-related illnesses is still a major health concern, especially since vast amounts of asbestos-containing products were manufactured, processed, and used over the past century (Kamp, 2009) and its long latency period that averages between 35 to 40 years from initial exposure to the first appearing symptoms (Ross, 1984). The toxic effects of asbestos inhalation depend on the cumulative dose, the time of initial exposure, and both the physical and chemical properties common to the different asbestos fibres (Kamp, 2009).
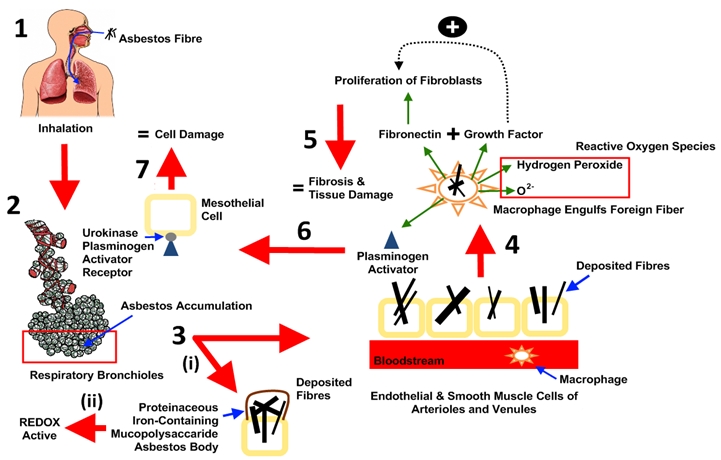
To gain an appreciation of the mechanism of damage induced by asbestos fibres, the four quintessential pillars of toxicity assessment must be explored, namely: Absorption, distribution, metabolism, and excretion. Since the main route of exposure is inhalation, the lungs, which comprise the tracheobronchial and the pulmonary or alveolar region, are the focal point where asbestos fibres initiate toxicity (Bignon et al. 1978). Histologically, fibrosis begins around the respiratory bronchioles in the lower lobes, where asbestos tends to accumulate and get absorbed upon inhalation (Gefter et al., 1992). In an acute study conducted by Brody et al. (1989), it was reported that lung damage to endothelial and smooth muscle cells of the arterioles and venules near the bifurcations surrounding this region, occurred within 19 to 72 hours subsequent to exposure in rats treated with four million fibres per m3 of chyrsotile. Fibres deposited in the bifurcations of alveolar ducts induce alveolar macrophages to attack at the site of deposition (Kamp, 2009). When the macrophage engulfs the foreign fibre, it produces cytokines such as fibronectin – a glycoprotein that recruits and initiates the proliferation of fibroblasts. In addition, the macrophages release excessive amounts of growth factors which act synergistically to produce an event cascade that leads to proliferation of more fibroblasts and collagen fibre deposition (Maxim & McConnell, 2001). As a result, fibrosis and scar accumulate which irreversibly alter pulmonary structure and function. Accumulating evidence also suggests that once macrophages engulf asbestos fibres, it forms and releases reactive oxygen species, such as hydrogen peroxide and superoxide anion O2- (Kamp & Weitzman, 1999), as well as plasminogen activators which, in turn, activate urokinase-type plasminogen activator receptors on the surface of human mesothelial cells (Hamilton et al., 1999). Once activated, damage to lung tissue occurs which further contributes to the future remodelling of the lung tissue attacked (Hamilton et al., 1999). Overtime, asbestos fibres deposited in the lungs are encapsulated by proteinaceous iron-containing mucopolysaccaride, forming redox-activeasbestos bodies (Kamp, 2009). A histological characteristic of asbestosis is the presence of pulmonary fibrosis in association with numerous asbestos bodies (Gee & Morgan, 1995). A schematic depicting these cellular events is illustrated in Figure 3.
Unlike other carcinogens, asbestos cannot be distributed through the bloodstream. If penetration does occur in lung or gastrointestinal tissue, it is extremely limited (Cook, 1983). Consequently, asbestos cannot be metabolized – once inhaled, it is retained for life. Seshan (1983) reported that simulated gastric juices can alter the physical and chemical properties of chrysotile and crocidolite fibres, but it does not stimulate metabolism. The low pH inside alveolar macrophages has also been speculated to weaken inhaled chyrsotile fibres and facilitate destruction (Finley et al., 2008). Finally, nearly all ingested fibres are excreted in the feces within 48 hours.
The idea linking asbestos genotoxic and carcinogenic potential through ingestion is a controversial supposition based on the notion that ingested fibres that come in contact with epithelial cells lining the lumen will penetrate the gastrointestinal tract and be distributed throughout the body. However, in a chronic study conducted by Chouroulinkov & Truhaut (1989), rats that were exposed to high oral doses of chrysotile and a mixture of chrysotile and crocidolite over a two year period showed no toxic or carcinogenic effects. Moreover, rats that were administered 0.5 to 50 mg chrysotile for 14 months, in another study showed significant increases in thymidine incorporation in the gastrointestinal tract (RAIS, 1995), suggesting that ingestion may interfere with DNA metabolism. As of yet, no oral studies have been conducted on humans, largely because of the difficulty in controlling external factors that influence the exposed tissue. Overall, to be a significant health concern, asbestos must either be inhaled, or to a lesser extent, ingested.
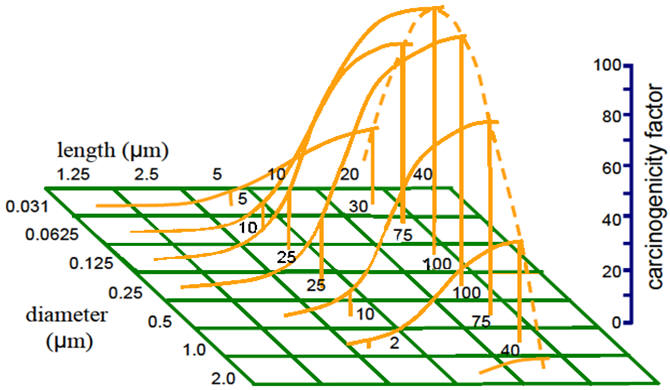
Size and shape of fibres play an important role in the potency and pathogenesis of asbestos-related illnesses. In a chronic study conducted by Layard et al. (1981), the occurrence of neoplastic tissue in rats correlated well with the dimensional distribution of asbestos fibres. They discovered that fibres ≤1.5 μm in diameter and >8 μm in length yielded the highest incidence of soft tissue sarcoma of the pleural cavity. If a fibre is longer than the macrophage diameter, it has the potential to reside in the lung for longer periods of time (Maxim & McConnell, 2001). In contrast, fibre ≤8 μm are often inactivated by phagocytosis (Layard et al., 1981) and removed from the lung either directly to the conducting airways or via lymphatics to regional lymph nodes (Maxim & McConnell, 2001). Pott (1978) provides a hypothetical view of how the carcinogenicity of asbestos varies in terms of length and diameter (Figure 4).
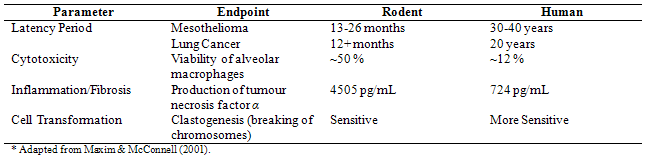
To date, all levels of asbestos exposure studies have demonstrated clinical effects. However, most literature published on dosimetry of the carcinogenic potency of asbestos is based on rats, as opposed to humans. Consequently, there are still many difficulties outlining precise doses and of what type of fibre will initiate pathogenic events in lung tissues. Although the risk of asbestos-related diseases is dose-response related (IPSC, 1998), a generalized dose-response curve is nonexistent for asbestos in human studies due to, and not limited to, time constraints (long latency period), extent of control, nature of the material, and external influences such as cigarette smoking (Kava, 2007). Dosimetry models that are available from cohort studies indicate that, on a normalized basis, fibre deposition and clearance rates are lower in humans than rats; this is because humans are generally much more sensitive (by a factor of 102 to 103) to fibres than rodents (Maxim & McConnell, 2001). Consequently, the use of rats as a model species to assess asbestos toxicity is limited. An interspecies comparison of various asbestos exposure parameters in rodents and humans are shown in Table 3.