Rickettsia prowazekii is the pathogenic bacterium responsible for epidemic typhus fever, an infectious disease that has plagued humans ever since it was first recorded in Europe nearly 1000 years ago (Szybalski, 1999). More recently, R. prowazekii is estimated to have infected nearly 30 million humans following the First and Second World War (Andersson et al., 1998), and continues to pose a major health threat in certain parts of the world, despite the availability of antibiotics. R. prowazekii is typically transmitted to humans by the body louse – an obligate ectoparasitic arthropod (Figure 1) (Baxter, 1996). In its vegetative state, the pleomorphic R. prowazekii is 0.3 μm to 0.5 μm by 0.8 μm to 2.0 μm in size (Walker, 1998), which makes this species too small to be clearly seen under an ordinary light microscope. R. prowazekii belongs to a large group of Gram-negative bacteria known as α-proteobacteria that multiply in eukaryotic cells. This suggests that they can only grow and reproduce within the living cells of their host. Although classified as Gram-negative bacteria, R. prowazekii are poorly stained by the Gram method and are better visualized using the Giemsa or Giménez stains (Raoult et al., 2004).

Intracellular R. prowazekii often appear as short, paired or single, lanceolate-to-ovoid cocco-bacilli (Raoult et al., 2004). To undergo cellular respiration, this species requires oxygen as a terminal electron acceptor. Its envelope consists of three major layers: an in-innermost cytoplasmic membrane, a thin electron-dense rigid cell wall containing peptidoglycan, and an outer layer, which contains lipopolysaccharide endotoxin (Yu & Walker, 2005) and immunodominant surface-exposed proteins that provide structure or potentially contribute to host cell adhesion or other host cell interactions (Figure 2) (Raoult et al., 2004). Cells of R. prowazekii also possess intracytoplasmic invaginations of the plasma membrane – a morphological feature that resembles cristae found in the mitochondria. Unlike the mitochondria, however, members of this species possess a flagellum for motility – a feature not common among the family Rickettsiaceae (Yu & Walker, 2005).

Since R. prowazekii requires an intracellular habitat, cells must be co-cultivated in tissue culture or yolk sac of developing chicken embryos (Figure 3); hence, a characteristic colonial morphology is inexistent. Under poor nutritional conditions, cells of R. prowazekii grow into long, 4 μm filaments and cease to multiply via transverse binary fission (Yu & Walker, 2005). However, when proper conditions are restored, they immediately divide into their ordinary short rod form and engage in extensive movements until released by the disruption of massively infected cells (Tritz, 2000).
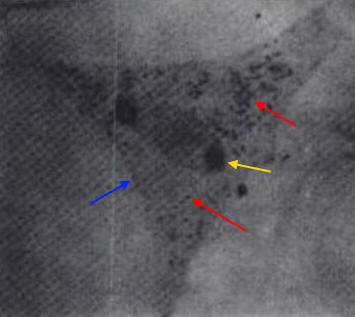
The nutritional requirements of R. prowazekii, outside their host cell, are not known (Yu & Walker, 2005). What is known is that R. prowazekii acquires most of its amino acids from its host cell because only the genes associated with the biosynthesis of lysine and serine are present in its DNA (Yu & Walker, 2005). In addition, maximal growth of this species can only occur if there is a sufficient amount of host cell proline and serine or glycine (Yu & Walker, 2005). This implies that R. prowazekii has a highly permeable cytoplasmic membrane. Likewise, glycolytic intermediates and products, such as acetyl-CoA, must also be obtained by its host. Similar to the mitochondrion found in eukaryotic cells, this species produces its own physiological energy supply of ATP via the enzyme ATPase (Yu & Walker, 2005). This reason, along with the evidence suggesting that R. prowazekii shares a phylogenetic relationship with the mitochondria, is why some biologists speculate that this species may have given rise to the modern-day mitochondria. Altogether, to be able to identify this species for diagnostic purposes, samples containing bacterial cells must be cultivated in tissue culture at 35°C. Once nutrients are depleted, longer, more filamentous-like cells with prominent vacuoles will develop (Figure 4). Infected tissues stained with Giemsa should show bluish-purple organisms, whereas tissue stained with Giménez should show bright red organisms, with a decolorized background stain of pale greenish blue (Yu & Walker, 2005). Due to the inability to grow R. prowazekii on ordinary microbiological media, such as nutrient broth, diagnosis is often difficult (Ge et al., 2004).
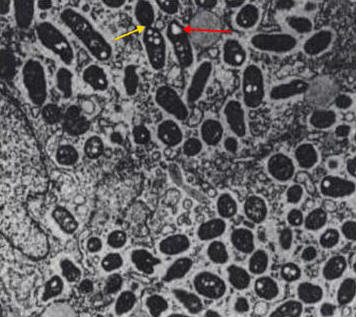
The entry of R. prowazekii into a human host cell (typically a phagocyte) involves three steps: attachment (via OmpA protein and a host cell receptor), internalization, and escape from the phagosome, prior to lysosomal fusion (Figure 5) (Yu & Walker, 2005). By escaping phagosomal digestion, this evasion mechanism allows the pathogen to reside in the cytoplasm and escape class II MHC presentation. However, the pathogenic mechanism used particularly by R. prowazekii as a means to escape phagosomal entrapment is unclear, but may involve potential virulence factors, such as phospholipase A (Winkler and Miller, 1982), antigenic surface proteins, hemolysin, type IV secretion systems, or hydrolase enzymes (Ge et al., 2004).
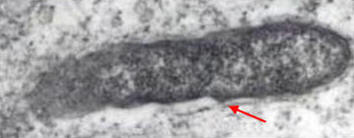
Body louse (Pediculus humanus humanus), the flying squirrel flea (Orchopeas howardii), and louse (Neohematopinus sciuropteri) are the most effective vectors in the spread of R. prowazekii-induced epidemic typhus fever. Once transmitted to the mammalian host through a bite via arthropod saliva, the bacteria are found principally in the endothelium of blood vessels, particularly in those of the brain, skin and heart (Tritz, 2000). This, in turn, causes hyperplasia of endothelial cells and localized thrombus formation, resulting in petechial rash, fever – which may reach 39°C – and terminal shock (Tritz, 2000). Finally, areas of poor hygiene, such as refugee camps during a major famine or natural disaster, are places where diseases such as typhus fever tend to flourish. In fact, due to recent outbreaks in parts of Europe, Africa, and South America, some experts support the notion that epidemic typhus is a reemerging public health problem (Ge et al., 2004).
References
Andersson, G.E., Zomorodipour, A., Andersson, J.O., Sicheritz-Pontén, T., Alsmark, C.M., Podowski, R.M., Näslund, K.A., Eriksson, A-S., Winkler, H.H., & Kurland, C.G. (1998). The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature, 396: 133-143.
Baxter, J.D. (1996). The Typhus Group. Clinical Dermatology, 14(3): 271-278.
Ge, H., Chuang, Y-Y.E., Zhao, S., Tong, M., Tsai, M-H., Temenak, J.J., Richards, A.L., & Ching, W.M. (2004). Comparative Genomics of Rickettsia prowazekii Madrid E and Breinl Strains. Journal of Bacteriology, 186(2): 556-565.
Raoult, D., Woodward, T., & Dumler, J.S. (2004). The history of epidemic typhus. Infectious Disease Clinics of North America,18(1): 127-140.
Szybalski, W. (1999). Maintenance of human-fed live lice in the laboratory and production of Weigl’s exanthematous typhus vaccine. In Maintenance of human, animal, and plant pathogen vectors, K. Maramorosch and F. Mahmood (eds.). Science Publishers Inc., Enfield, New Hampshire, 161–179.
Tritz, G. (2000). Rickettsia, Chlamydia, Mycoplasma. Retrieved February 16th, 2010, from: http://www.atsu.edu/faculty/chamberlain/Website/Lects/rickett.htm.
Walker, D.H. (1988). Pathology and pathogenesis of the vasculotropic rickettsioses, In: D.H. Walker (Ed.), Biology of Rickettsial Disease, Florida: CRC Press Inc. 115-138.
WHO: World Health Organization. (1976). Centers for Disease Control and Prevention – Public Health Image Library (CDC PHIL), I.D. # 5289.
Winkler, H.H., & Miller, E.T. (1982). Phospholipase A and the Interaction of Rickettsia prowazekii and Mouse Fibroblasts (L-929 Cells). Infection & Immunity, 38(1): 109-113.
Yu, X-J., & Walker, D.H. (2005). Genus I. Rickettsia da Rocha-Lima 1916. In Bergey’s Manual of Systematic Bacteriology, 2nd edn., vol. 2 (The Proteobacteria). 96-106. Edited by Brenner D.J., Krieg N.R., & Garrity, G.M., & Staley J. T. New York: Springer.