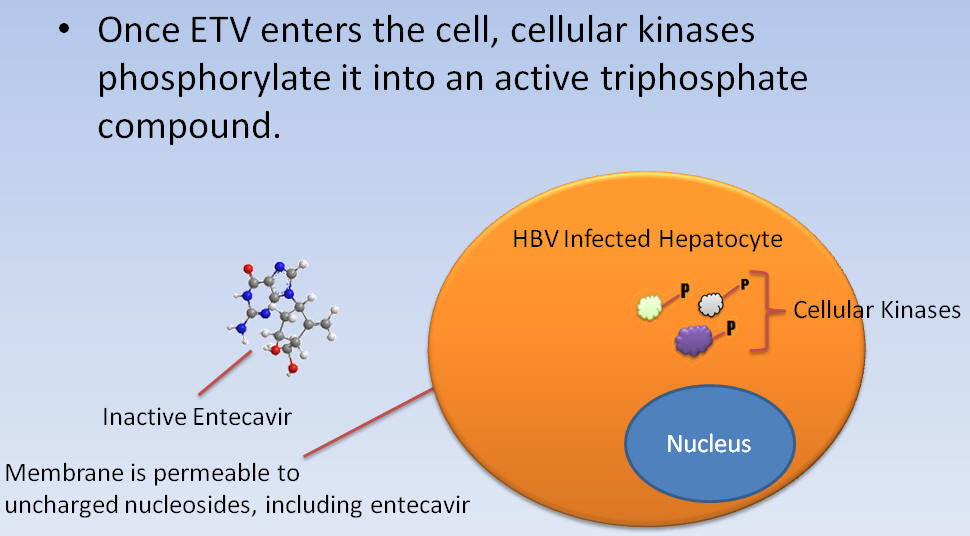
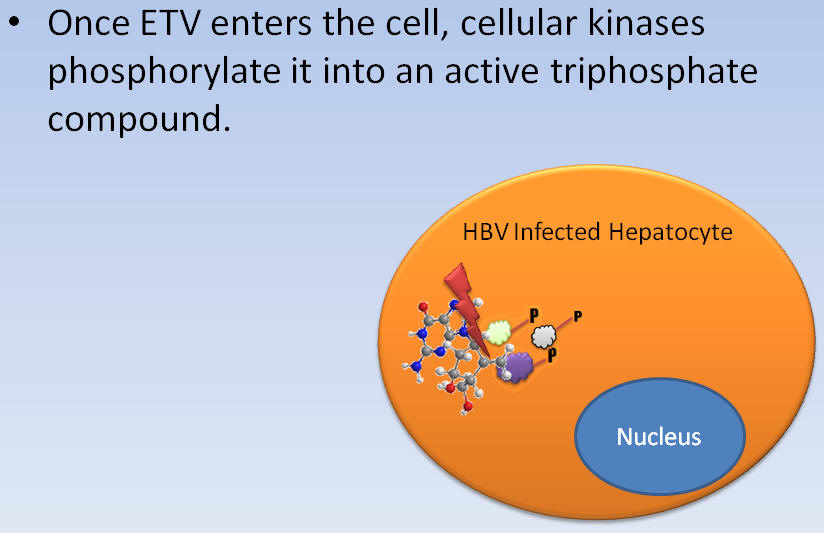
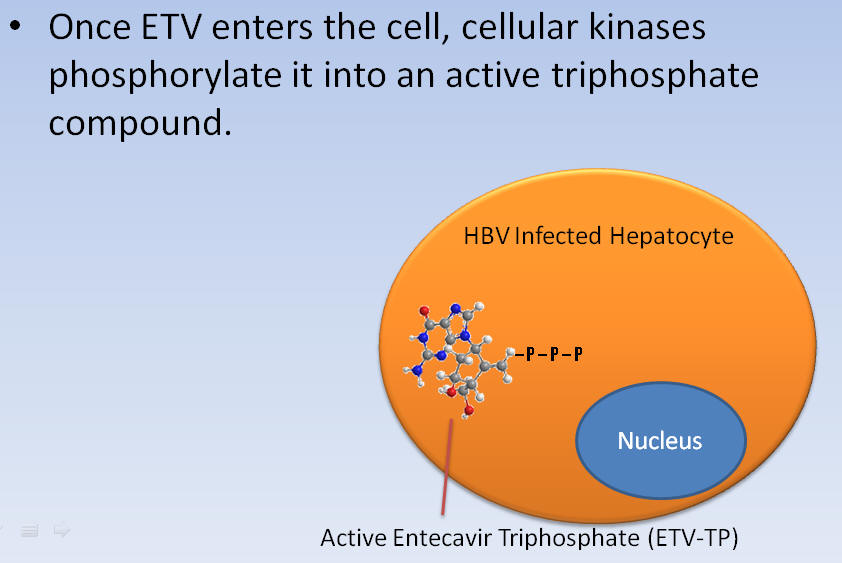
Once ETV enters the cell, cellular kinases phosphorylate it into an active triphosphate compound.
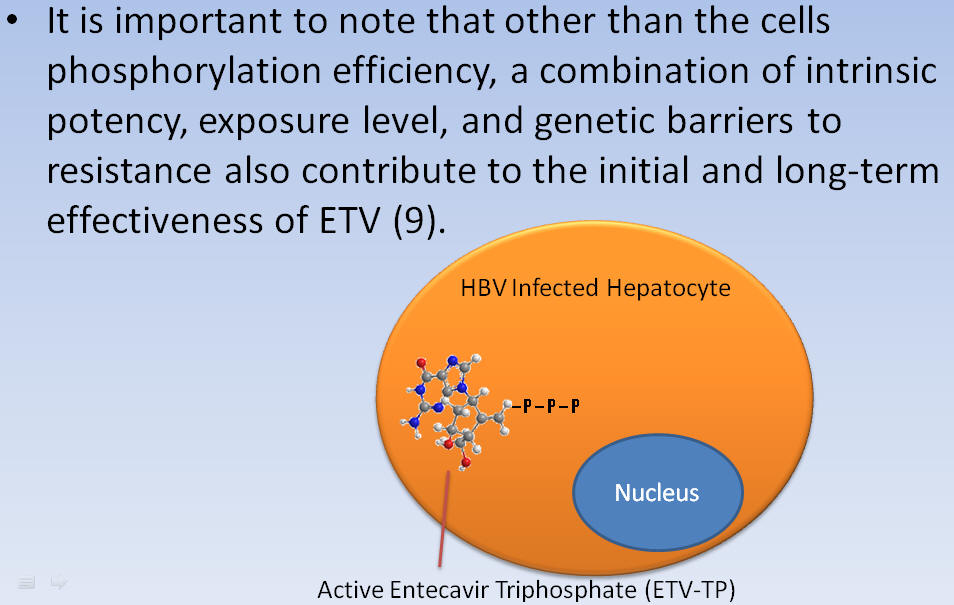
It is important to note that other than the cells phosphorylation efficiency, a combination of intrinsic potency, exposure level, and genetic barriers to resistance also contribute to the initial and long-term effectiveness of ETV.
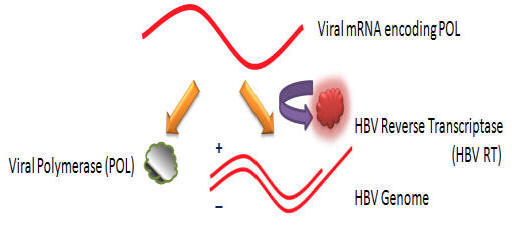
ETV inhibits the only known enzymatic target of the hepatitis B virus, the viral polymerase (Pol). The mRNA encoding POL serves as the template for synthesis of the hepatitis B virus genome via reverse transcriptase activity.
While cellular polymerases and reverse transcriptases use free hydroxyl groups of DNA or RNA as primers for DNA synthesis, the DNA polymerases of certain animal viruses, such as the hepatitis B virus, depend upon hydroxyl groups of amino acid residues within proteins (10). For instance, the primer for Pol DNA synthesis is a hydroxyl group of a tyrosine residue near the amino terminus of Pol.
ETV triphosphate (ETV-TP) displays activity against the synthetic activities of HBV polymerase:
- The unique protein-linked priming activity
- RNA-directed first-strand synthesis via reverse transcription
- Second-strand DNA-directed DNA synthesis (Not Shown)
Recall that the hepatitis B virus genome is partially single-stranded (minus) and partially double-stranded (plus).
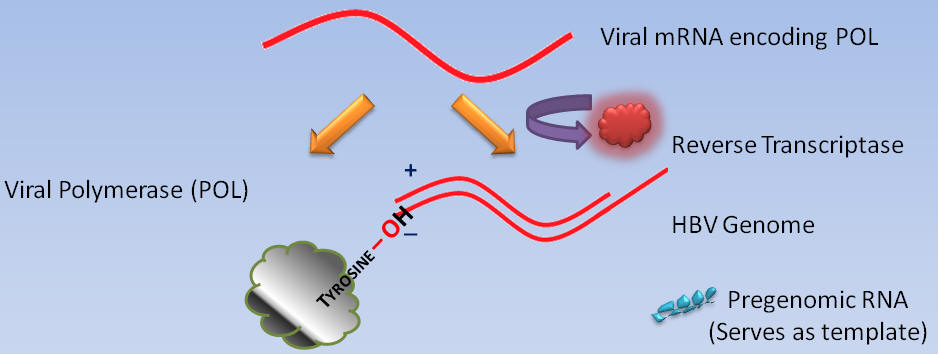
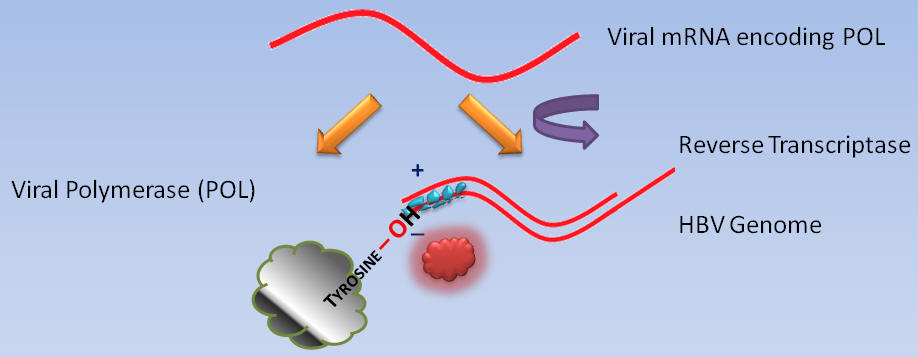
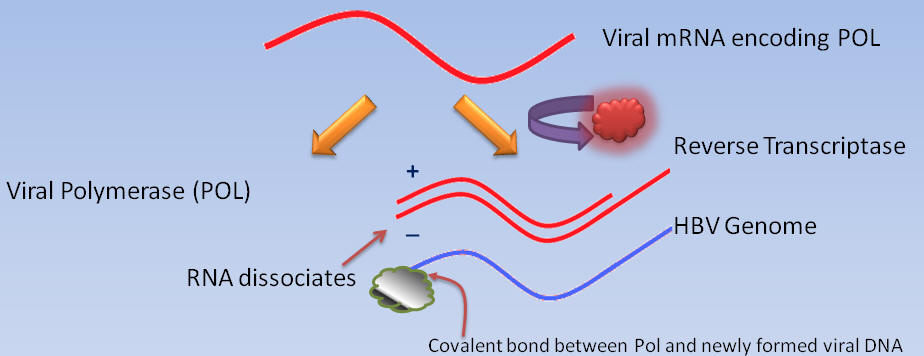
In order to understand the interaction of ETV with the hepatitis B virus at the molecular level, ETV triphosphate must be modeled in the catalytic site of the hepatitis B virus reverse transcriptase, as well as in the process of the hepatitis B virus DNA elongation.
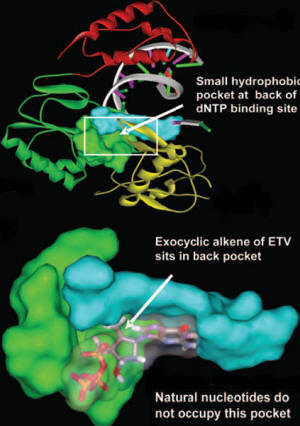
- Unlike conventional hepatitis B viral treatments, ETV is unique in having a D-configured cyclopentyl group.
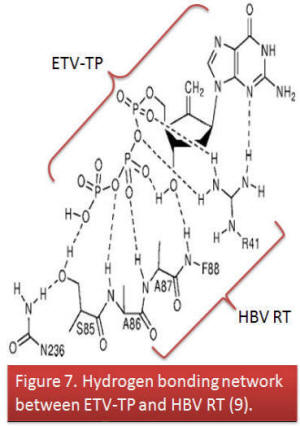
- During translation of viral genome, ETV fits directly into the hydrophobic pocket of hepatitis B virus reverse transcriptase at the back dNTP binding site via hydrogen bonds. Normal dGTP cannot wholly occupy the pocket during replication since it does not contain a cyclopentyl group like ETV. Recall that ETV is a guanosine nucleoside analogue (9).
- This property accounts for the higher affinity of ETV for the hepatitis B virus reverse transcriptase than that of dGTP, and higher potency compared to other drugs (9).
- The absence of this small pocket in human polymerase correlates with the relative inactivity of ETV during host DNA replication.
Since ETV possesses a potential 3’ hydroxyl group, it is important to establish the effect of ETV incorporation on HBV DNA chain extension.
The mechanism for chain termination by ETV likely involves incorporation and abortive extension of ETV-containing DNA. This can occur in any three instances (9,11):
- Initial docking into the dNTP binding site and addition of ETV onto the 3’ end of the growing DNA
- Upon addition of ETV in the +1 position, and
- Upon further addition of nucleotides when ETV is elongated to the +2 position.
When ETV enters HBV RT, its cyclopentyl group causes steric strain on the HBV DNA, thereby distorting and partially blocking the dNTP binding site. As a result, this prevents the binding of any new substrate and resulting in chain termination (11).